Stellate cell
Star-shaped neurons in the central nervous system
| Stellate cell | |
|---|---|
 Golgi stained cortical neurons A) Layer II/III pyramidal cell B) layer IV spiny stellate cell | |
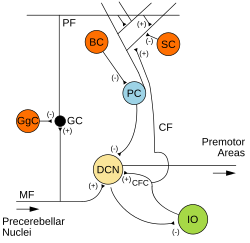 Microcircuitry of the cerebellum. Excitatory synapses are denoted by (+) and inhibitory synapses by (-). MF: Mossy fiber. DCN: Deep cerebellar nuclei. IO: Inferior olive. CF: Climbing fiber. GC: Granule cell. PF: Parallel fiber. PC: Purkinje cell. GgC: Golgi cell. SC: Stellate cell. BC: Basket cell. | |
| Identifiers | |
| NeuroLex ID | sao2046525601 |
| Anatomical terms of neuroanatomy [edit on Wikidata] | |
Stellate cells are neurons in the central nervous system, named for their star-like shape formed by dendritic processes radiating from the cell body. These cells play significant roles in various brain functions, including inhibition in the cerebellum and excitation in the cortex, and are involved in synaptic plasticity and neurovascular coupling.
Morphology
Stellate cells are characterized by their star-shaped dendritic trees. Dendrites can vary between neurons, with stellate cells being either spiny or aspinous. In contrast, pyramidal cells, which are also found in the cerebral cortex, are always spiny and pyramid-shaped. The classification of neurons often depends on the presence or absence of dendritic spines: those with spines are classified as spiny, while those without are classified as aspinous.
Types and locations
Cerebellar
Many stellate cells are GABAergic and are located in the molecular layer of the cerebellum.[1] Most common stellate cells are the inhibitory interneurons found within the upper half of the molecular layer in the cerebellum. These cells synapse onto the dendritic trees of Purkinje cells and send inhibitory signals.[2] Stellate cells are derived from dividing progenitor cells in the white matter of the postnatal cerebellum.
Cortical
Stellate neurons are also found in the cortex. Cortical spiny stellate cells are located in layer IVC of the primary visual cortex,[3] and in the somatosensory barrel cortex of mice and rats, glutamatergic (excitatory) spiny stellate cells are organized in the barrels of layer 4.[4] These cells receive excitatory synaptic fibers from the thalamus and process feed-forward excitation to layers 2/3 of the primary visual cortex to pyramidal cells. Cortical spiny stellate cells exhibit a 'regular' firing pattern.
Other locations
GABAergic aspinous stellate cells are also found in the somatosensory cortex. These cells can be immunohistochemically labeled with glutamic acid decarboxylase (GAD) due to their GABAergic activity, and they occasionally colocalize with neuropeptides.[5]
Development
Stellate and basket cells originate from the cerebellar ventricular zone (CVZ) along with Purkinje cells and Bergmann glia.[6]: 283 [7] These cells follow a similar pathway during migration, starting in the deep layer of the white matter, moving through the internal granular layer (IGL) and the Purkinje cell layer (PCL) until reaching the molecular layer.[6]: 284 In the molecular layer, stellate cells change orientation and positioning until they reach their final placement, guided by Bergmann glial cells.[8]
Function
Stellate cells receive Excitatory Post Synaptic Potentials (EPSCs) from parallel fibers. The characteristics of these EPSCs depend on the pattern and frequency of presynaptic activity, influencing the extent and duration of inhibition within the cerebellar cortex.[9] Synapses between parallel fibers and stellate cells exhibit plasticity, allowing for long-term changes in synaptic efficacy. This synaptic plasticity can occur at both parallel fiber-stellate cell synapses and parallel fiber-Purkinje cell synapses, suggesting a role in cerebellar motor learning.[10]
Neurovascular Coupling
Cerebellar stellate cells also play a crucial role in neurovascular coupling. Electrophysiological stimulation of single stellate cells is sufficient to release nitric oxide (NO) and induce dilation of blood vessels.[11]
See also
References
- ^ Rubenstein J, Rakic P (2013). Patterning and Cell Type Specification in the Developing CNS and PNS : Comprehensive Developmental Neuroscience. Elsevier Science. p. 215. ISBN 978-0-12-397265-1.[permanent dead link]
- ^ Chan-Palay V, Palay SL (1972-01-01). "The stellate cells of the rat's cerebellar cortex". Zeitschrift Fur Anatomie Und Entwicklungsgeschichte. 136 (2): 224–248. doi:10.1007/BF00519180. PMID 5042759. S2CID 8003308.
- ^ da Costa NM, Martin KA (February 2011). "How thalamus connects to spiny stellate cells in the cat's visual cortex". The Journal of Neuroscience. 31 (8): 2925–2937. doi:10.1523/JNEUROSCI.5961-10.2011. PMC 6623786. PMID 21414914.
- ^ Petersen CC (October 2007). "The functional organization of the barrel cortex". Neuron. 56 (2): 339–355. doi:10.1016/j.neuron.2007.09.017. PMID 17964250.
- ^ Conn PM (2008). Neuroscience in medicine (3rd ed.). Beaverton, OR: Humana Press. ISBN 978-1-60327-454-8.
- ^ a b Munz M, Ruthazer ES (2013). Comprehensive developmental neuroscience. Cellular migration and formation of neuronal connections (First ed.). Elsevier Science & Technology. p. 283. ISBN 978-0-12-397266-8.
- ^ "Cerebellar Ventricular Zone - Cellular Development, Function & Anatomy". LifeMap Sciences, Inc.
- ^ Rubenstein J (2013-05-06). Patterning and Cell Type Specification in the Developing CNS and PNS: Comprehensive Developmental Neuroscience. Elsevier Science & Technology. ISBN 978-0-12-397348-1.
- ^ Rancillac A, Barbara JG (May 2005). "Frequency-dependent recruitment of inhibition mediated by stellate cells in the rat cerebellar cortex". Journal of Neuroscience Research. 80 (3): 414–423. doi:10.1002/jnr.20473. PMID 15789412.
We found that single intense stimulations mostly produce individual SC EPSCs with large amplitude and variable latencies, but they often fail. Increasing the stimulation frequency above 60 Hz reduces failures but only slightly increases the mean amplitude. Reducing failures at PF-SC synapses increases the number of SC EPSCs per stimulation but also only slightly increases the mean amplitude. Brief bursts of presynaptic activity temporarily depress synaptic transmission due to endocannabinoid release, serving as a feedback mechanism.
- ^ Rancillac A, Crépel F (February 2004). "Synapses between parallel fibres and stellate cells express long-term changes in synaptic efficacy in rat cerebellum". The Journal of Physiology. 554 (Pt 3): 707–720. doi:10.1113/jphysiol.2003.055871. PMC 1664787. PMID 14617674.
We show that long-term potentiation (LTP) and long-term depression (LTD) were induced at these synapses by a low frequency stimulation protocol (2 Hz for 60 s) and that pairing this low frequency stimulation protocol with postsynaptic depolarization induced a marked shift of synaptic plasticity in favour of LTP.
- ^ Rancillac A, Rossier J, Guille M, Tong XK, Geoffroy H, Amatore C, et al. (June 2006). "Glutamatergic Control of Microvascular Tone by Distinct GABA Neurons in the Cerebellum". The Journal of Neuroscience. 26 (26): 6997–7006. doi:10.1523/JNEUROSCI.5515-05.2006. PMC 6673912. PMID 16807329.
Cerebellar stellate and Purkinje cells dilate and constrict, respectively, neighboring microvessels. This highlights the specialized functions of different neuron types in regulating cerebral blood flow, emphasizing the complex interplay between various neurons in maintaining neurovascular balance.
External links
- NIF Search - Stellate Cell via the Neuroscience Information Framework
- v
- t
- e
Anatomy of the cerebellum
| Lobes | |
|---|---|
| Medial/lateral |
|
| Deep cerebellar nuclei | |
|---|---|
| Cerebellar cortex |
|
| Internal |
|
|---|---|
| Peduncles |










